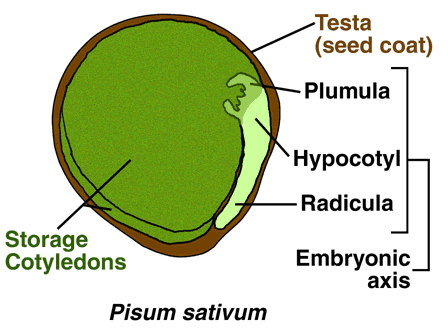
Seed Germination and and Dormancy are Regulated by Light, Temperature and Plant Hormones
Tobacco seed germination and class I ß-1,3-glucanase (ßGlu I) induction regulated by plant hormones
ABA-sensitive ßGlu expression in the endosperm prior to endosperm rupture is widespread among Solaneceous seeds
Abscisic acid (ABA)
Gibberellins (GA)
Light / phytochrome (Pfr)
Temperature
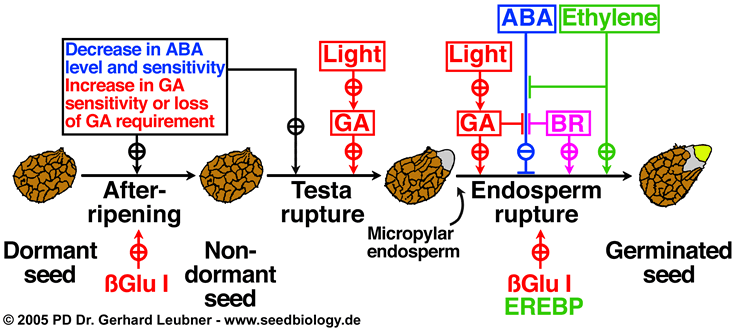
Hormonal interactions during seed dormancy release and germination
Table: Seed dormancy and germination of Arabidopsis hormone mutants
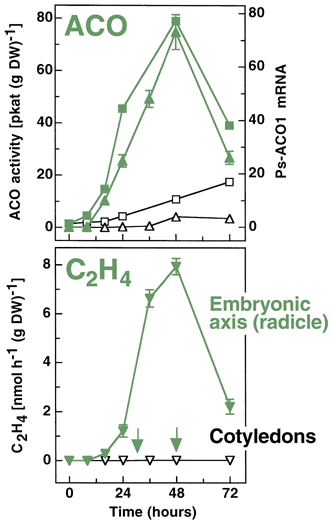
Ethylene and pea seed germination
Ethylene-Responsive Element Binding Proteins (EREBP)
Brassinosteroids and gibberellins promote tobacco seed germination by distinct pathways
|
Class I ß-1,3-glucanase (ßGlu I) induction is tightly linked with altered endosperm rupture in response to physiological factors known to affect the incidence and timing of tobacco seed germination
- Photodormant tobacco seeds do not germinate in darkness, but treatment with a red-light pulse or with gibberellin (GA) is sufficient to release photodormancy, induce testa rupture, ßGlu I induction and subsequent endosperm rupture in the dark (Leubner-Metzger et al., 1996, Leubner-Metzger 2001, Leubner-Metzger 2002). Release of photodormancy is blocked in antisense-ßGlu I seeds (Leubner-Metzger and Meins, 2001).
- ABA delays endosperm rupture and inhibits ßGlu I induction in a concentration-dependent manner, but does not affect photodormancy or testa rupture. Treatment of tobacco seeds with 10 µM ABA results in the formation of a novel structure, consisting of the enlarging radicle with a sheath of greatly elongated endosperm tissue (Leubner-Metzger et al., 1995).
- Ethylene promotes endosperm rupture and ßGlu I induction, but does not affect photodormancy or testa rupture. Ethylene-responsive element binding proteins (EREBPs) exhibit a novel pattern of expression during seed germination (Leubner-Metzger et al., 1998).
- Sense-transformation with a chimeric ABA-inducible ßGlu I transgene resulted in over-expression of ßGlu I in the seed covering layers and promoted endosperm rupture of mature seeds and of ABA-treated after-ripened seeds. In reciprocal crosses promotion of endosperm rupture only occurred if the mother plant was an over-expresser. Sense-transformation directly shows that ßGlu I has a causal role in endosperm rupture (Leubner-Metzger and Meins, 2000; Leubner-Metzger, 2002; Manz et al., 2005).
- Treatments promoting tobacco seed germination promote ßGlu I induction (e.g. light, GA, ethylene); Whereas treatments inhibiting germination inhibit ßGlu I induction (e.g.darkness, ABA, osmotica) (Reviews: Leubner-Metzger and Meins, 1999; Leubner-Metzger, 2003).
- Tobacco seed after-ripening is correlated with ßGlu I expression and the promotion of testa rupture (Leubner-Metzger, 2005). The after-ripening-mediated promotion of testa rupture is blocked in antisense-ßGlu I seeds (Leubner-Metzger and Meins, 2001) and ßGlu I over-expression can replace the after-ripening effect on the promotion of testa rupture (Leubner-Metzger and Meins, 2000; Leubner-Metzger, 2002).
|
Summary of the hormonal regulation of tobacco seed dormancy release, seed germination and ßGlu I expression. Note that tobacco seed germination is a two-step process with testa rupture followed by endosperm rupture.
Review "Plant hormone interactions during seed dormancy release and germination"by Kucera, Cohn, Leubner-Metzger, Seed Science Research 15: 281-307 (2005)
|
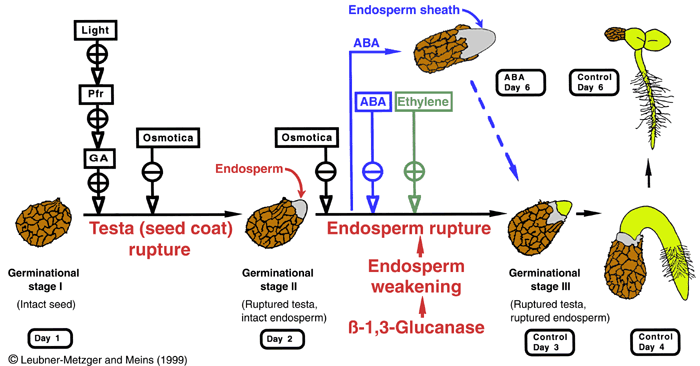
Figure 3.1: Working model for tobacco seed germination. Rupture of the testa and rupture of the endosperm are separate events in Nicotiana tabacum. Class I ß-1,3-glucanase (ßGLU I) accumulates just prior to endosperm rupture and is proposed to promote radicle protrusion by weakening of the endosperm. Plant hormones and environmental factors alter the germination process and in strict correlation with this either promote (+) or inhibit (-) ßGLU I induction. GA = gibberellin(s); ABA = abscisic acid; Pfr = Phytochrome. The model summarizes results from Leubner-Metzger et al. (1995, 1996, 1998).
© Review "Functions and Regulation of Plant ß-1,3-Glucanases (PR-2)" by Leubner-Metzger and Meins (1999)
© Review "Functions and regulation of ß-1,3-glucanases during seed germination, dormancy release and after-ripening"
by Leubner-Metzger, Seed Science Research 13: 17-34 (2003)
|
Hormonal interactions during seed dormancy release and germination
Review "Plant hormone interactions during seed dormancy release and germination" by Kucera, Cohn, Leubner-Metzger, Seed Science Research 15: 281-307 (2005)
Table 1: Seed dormancy and germination of Arabidopsis hormone mutants
Figure 2 : Hormonal interactions during seed dormancy release and germination
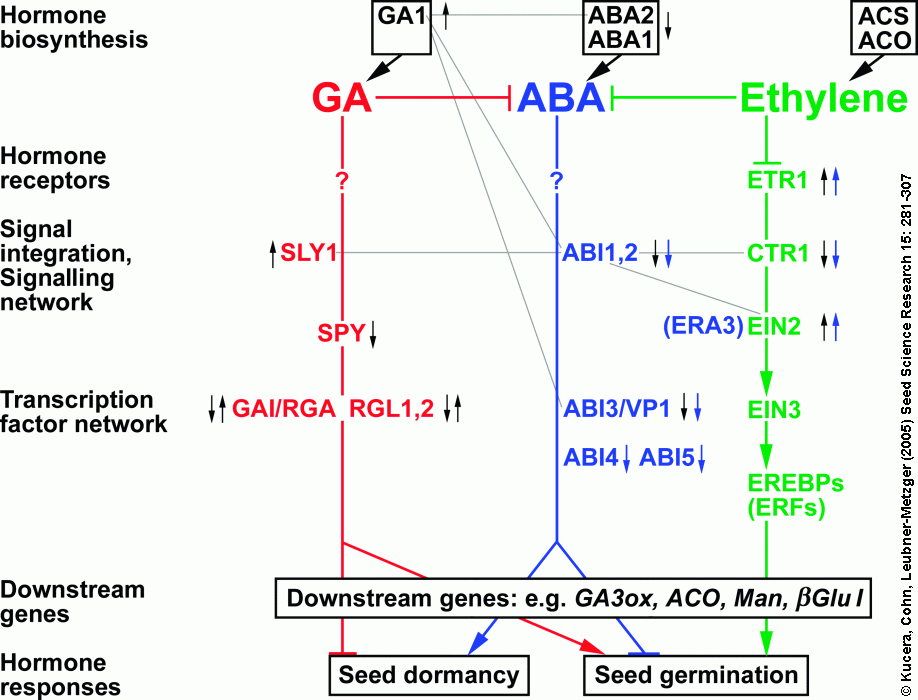
Figure 2. Schematic representation of the interactions between the gibberellin (GA), absisic acid (ABA) and ethylene signaling pathways in the regulation of seed dormancy and germination. The model is mainly based on Arabidopsis hormone mutant analyses, the positions of some components are speculative, and details are explained in the text. Promotion or inhibition is indicated by thick arrows and blocks, respectively. Interactions based on extragenic suppressor or enhancer screens are indicated by thin grey lines. Small black arrows indicate enhancement (up-arrow) or reduction (down-arrow) of seed dormancy and small blue arrows indicate enhancement or reduction of seed ABA sensitivity upon mutation of the corresponding protein. Corresponding hormone mutants of Arabidopsis thaliana: aba1, aba2 = ABA-deficient1,2; abi1 to abi5 = ABA-insensitive1 to ABA-insensitive5; ctr1 = constitutive triple response1; ein2, ein3 = ethylene insensitive2, 3; era3 = enhanced response to ABA3; gai = GA-insensitive; sly1 = sleepy1; spy = spindly; rga = repressor-of-ga1-3; rgl 1, rgl2 = rga-like1, 2. Other abbreviations: ACO = ACC oxidase; ACS = ACC synthase; EREBP = ethylene responsive element binding protein; ERF = ethylene responsive factor; GA3ox = GA 3-oxidase; Man = mannanase; ßGlu I = class I ß-1,3-glucanase; vp1 = viviparous1 (maize mutant). From:
Review "Plant hormone interactions during seed dormancy release and germination" by Kucera, Cohn, Leubner-Metzger, Seed Science Research 15: 281-307 (2005)
|
pea |
| |
|
 |
| |
|
|
|

|
|