The Plant Cell 21: 3803-3822 (2009)
Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: a comparative approach using Lepidium sativum and Arabidopsis thaliana [W][OA]
Warwick Horticulture Research International (HRI), Warwick University, Wellesbourne, Warwick CV35 9EF, United Kingdom (Ka.M., C.S.C.C., J.R.L., W.E.F.-S.)
Palacky University and Institute of Experimental Botany Academy of Sciences of the Czech Republic, Laboratory of Growth Regulators, CZ-78371 Olomouc, Czech Republic (V.T., M.S.)
Université Pierre et Marie Curie-Paris 6, Germination et Dormance des Semences, UR5, Site d'Ivry, F-75005 Paris, France (F.C.)
Received July 23, 2009; Returned for revision October 12, 2009; Accepted November 17, 2009; Published December 18, 2009
www.plantcell.org/cgi/doi/10.1105/tpc.109.070201
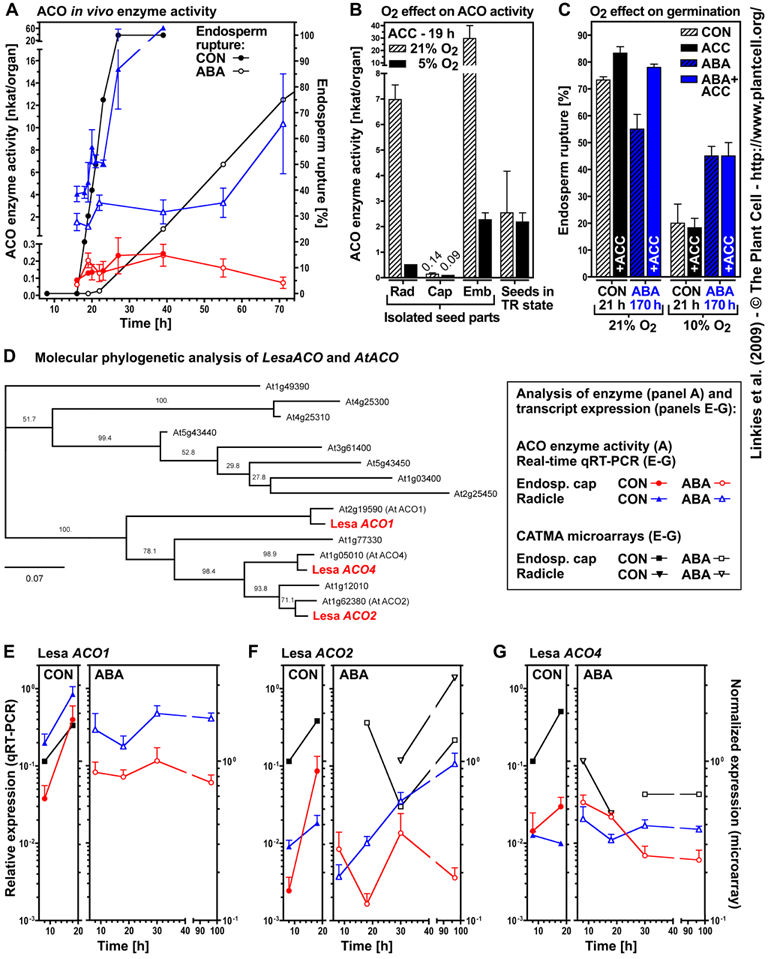
Figure 7. Analysis of ACC oxidase (ACO) enzyme activity and transcript expression in specific seed tissues of Lepidium sativum during germination.
The framed box shows symbols used in (A) and (E) to (G). Mean values ±SE ([A] to [C]) or +SE (qRT-PCR; [E] to [G]) are presented for three ([A] to [C]) or four ([E] to [G]) biologically independent samples each with 50 seeds (C), 100 to 200 seed parts ([A] and [B]) or 1000 endosperm caps or 100 radicles ([E] to [G]) from seeds with ruptured testa but intact endosperm ([A], [B], and [E] to [G]). L. sativum FR14 ([A] to [D]) or FR1 ([E] to [G]) seeds were incubated at 24°C in continuous light. Note that the qRT-PCR and the microarrays were performed as independent experiments.
(A) Time course of in vivo ACO enzyme activities in endosperm caps and radicles of seeds incubated without (CON) and with 10 µM ABA; for comparison, the kinetics of endosperm rupture are presented. In vivo ACO enzyme activities of endosperm caps and radicles dissected from seeds at different time points were measured by subsequent organ incubation in medium with ACC (plus ABA for the ABA series).
(B) The effect of ambient and reduced oxygen atmospheres on the in vivo ACO enzyme activities of isolated seed parts compared with intact seeds. Note that only seeds in the testa rupture (TR) state were selected for the measurements (i.e., seeds with intact endosperm). No wound-induced ethylene production by the radicle/embryo was evident.
(C) The effect of oxygen on the promotion of endosperm rupture by ACC and the reversion of the ABA inhibition of endosperm rupture by ACC.
(D) Molecular phylogenetic analysis of Lepidium ACO and Arabidopsis ACO cDNA sequences. The bar (0.07) defines the number of substitutions per 100 amino acids. Lepidium ACO cDNA sequences and comparisons to the Arabidopsis orthologs are presented in Supplemental Figure 3 online.
(E) to (G) LesaACO transcript expression pattern determined by qRT-PCR in endosperm cap and radicle during incubation on medium without (CON) or with 10 µM ABA added. Relative deltadeltaCt expression values based on the comparison with validated constitutive transcripts are presented.
Synopsis: Tissue weakening of the endosperm is required to allow radicle protrusion during seed germination. Cross-species work including tissue-specific transcriptome analysis and biomechanical measurement of endosperm weakening provided a new mechanistic model that explains how ethylene promotes seed germination and counteracts the inhibition of endosperm cap weakening by abscisic acid.
| Article in PDF format (1.8 MB) Supplemental data file (1.8 MB) Abstract of Plant Cell 2009 |
|
|
|
The Seed Biology Place |
Webdesign Gerhard Leubner 2000 |