Plant Physiology 160: 1551-1566 (2012)
Distinct cell wall architectures in seed endosperms in representatives of the Brassicaceae and Solanaceae [W][OA]
Centre for Plant Sciences, Faculty of Biological Sciences, University of Leeds, Leeds LS2 9JT, UK (KL, PK)
Wageningen Seed Lab, Laboratory of Plant Physiology, Wageningen University, Droevendaalsesteeg 1, 6708 PB, Wageningen, The Netherlands (BD, LB)
Department of Molecular Plant Physiology, Utrecht University, 3584 CH Utrecht, The Netherlands (BD, LB)
University of Freiburg, Faculty of Biology, Institute for Biology II, Botany/Plant Physiology, D-79104 Freiburg, Germany (TS*, GLM*)
ARC Centre of Excellence in Plant Cell Walls, School of Botany, University of Melbourne, Parkville, Victoria 3010, Australia (BD, LB)
* Current Address: School of Biological Sciences, Royal Holloway, University of London, Bourne Building 3-30, Egham, Surrey, TW20 0EX, UK
Received July 13, 2012; Accepted September 4, 2012; Published September 6, 2012.
DOI:10.1104/pp.112.203661
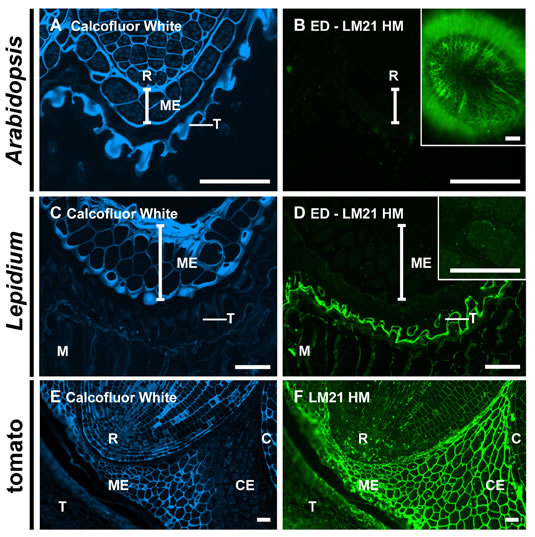
Supplemental Figure S2. Immunodetection of heteromannans in enzymatic deconstructed sections of Arabidopsis and Lepidium seeds and in untreated section of tomato seeds.
A, and B, LM21 heteromannan (HM) is not detectable in Arabidopsis enzyme deconstructed (ED) 3 h-imbibed seed sections but is weakly detectable in embryo cell walls of 3 h-imbibed Lepidium seeds (D, inset image).
B, LM21 HM is detectable in the mucilage of intact Arabidopsis seeds (inset image).
C and D, LM21 heteromannan is abundant and the mucilage (M) and at the testa surface (T) of Lepidium seeds.
E, Calcofluor White labelling of 3 h-imbibed tomato seed sections revealed tissue level asymmetry deliniating micropylar (ME) and chalazal endosperm (CE) regions of the endosperm.
F, LM21 labelling indicated abundant heteromannan in endosperm and embryo cell walls.
Vertical bars indicate extent of ME in Arabidopsis and Lepidium sections. (R) = radicle apex, (C) = cotyledons. Scale bars = 50 μm.
| Article in PDF format (1.5 MB) Supplementary data file (2 MB) |
|
|
|
The Seed Biology Place |
Webdesign Gerhard Leubner 2000 |