Plant Physiology 160: 1551-1566 (2012)
Distinct cell wall architectures in seed endosperms in representatives of the Brassicaceae and Solanaceae [W][OA]
Centre for Plant Sciences, Faculty of Biological Sciences, University of Leeds, Leeds LS2 9JT, UK (KL, PK)
Wageningen Seed Lab, Laboratory of Plant Physiology, Wageningen University, Droevendaalsesteeg 1, 6708 PB, Wageningen, The Netherlands (BD, LB)
Department of Molecular Plant Physiology, Utrecht University, 3584 CH Utrecht, The Netherlands (BD, LB)
University of Freiburg, Faculty of Biology, Institute for Biology II, Botany/Plant Physiology, D-79104 Freiburg, Germany (TS*, GLM*)
ARC Centre of Excellence in Plant Cell Walls, School of Botany, University of Melbourne, Parkville, Victoria 3010, Australia (BD, LB)
* Current Address: School of Biological Sciences, Royal Holloway, University of London, Bourne Building 3-30, Egham, Surrey, TW20 0EX, UK
Received July 13, 2012; Accepted September 4, 2012; Published September 6, 2012.
DOI:10.1104/pp.112.203661
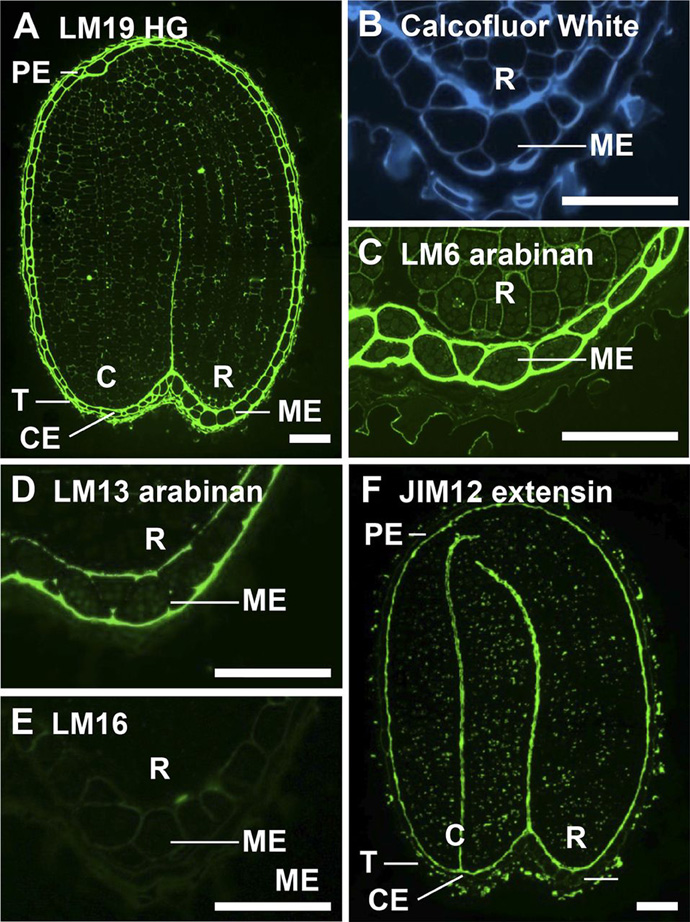
Figure 3. In situ localization of pectic polysaccharides and extensin in medial longitudinal sections of 3-h-imbibed Arabidopsis seeds.
A, Immunolabeling with LM19 revealed the abundance of unesterified HG in the endosperm.
B, Calcofluor White labeled all cell walls. Labeling with anti-arabinan probes revealed the spatial heterogeneity of arabinan in the endosperm.
C, LM6 arabinan was uniformly distributed through endosperm cell walls.
D, LM13 bound to the outer walls of the endosperm and weakly to transverse endosperm walls.
E, The LM16 epitope was weakly detectable in endosperm walls.
F, JIM12 labeling of extensin indicated restricted occurrence at the embryo surface/inner face of the endosperm and testa surface. C, Cotyledons; R, radicle; T, testa. Bars = 50 mm.
| Article in PDF format (1.5 MB) Supplementary data file (2 MB) |
|
|
|
The Seed Biology Place |
Webdesign Gerhard Leubner 2000 |