Tansley Review - New Phytologist 186: 817-831 (2010)
The evolution of seeds
Biological Sciences Department, California Polytechnic State University, San Luis Obispo, 7 CA 93401, USA (C.A.K.)
*Corresponding authors: C.A.K., G.L.-M.
Received December 21, 2009; accepted February 23, 2010; published online April 12, 2010
DOI 10.1111/j.1469-8137.2010.03249.x
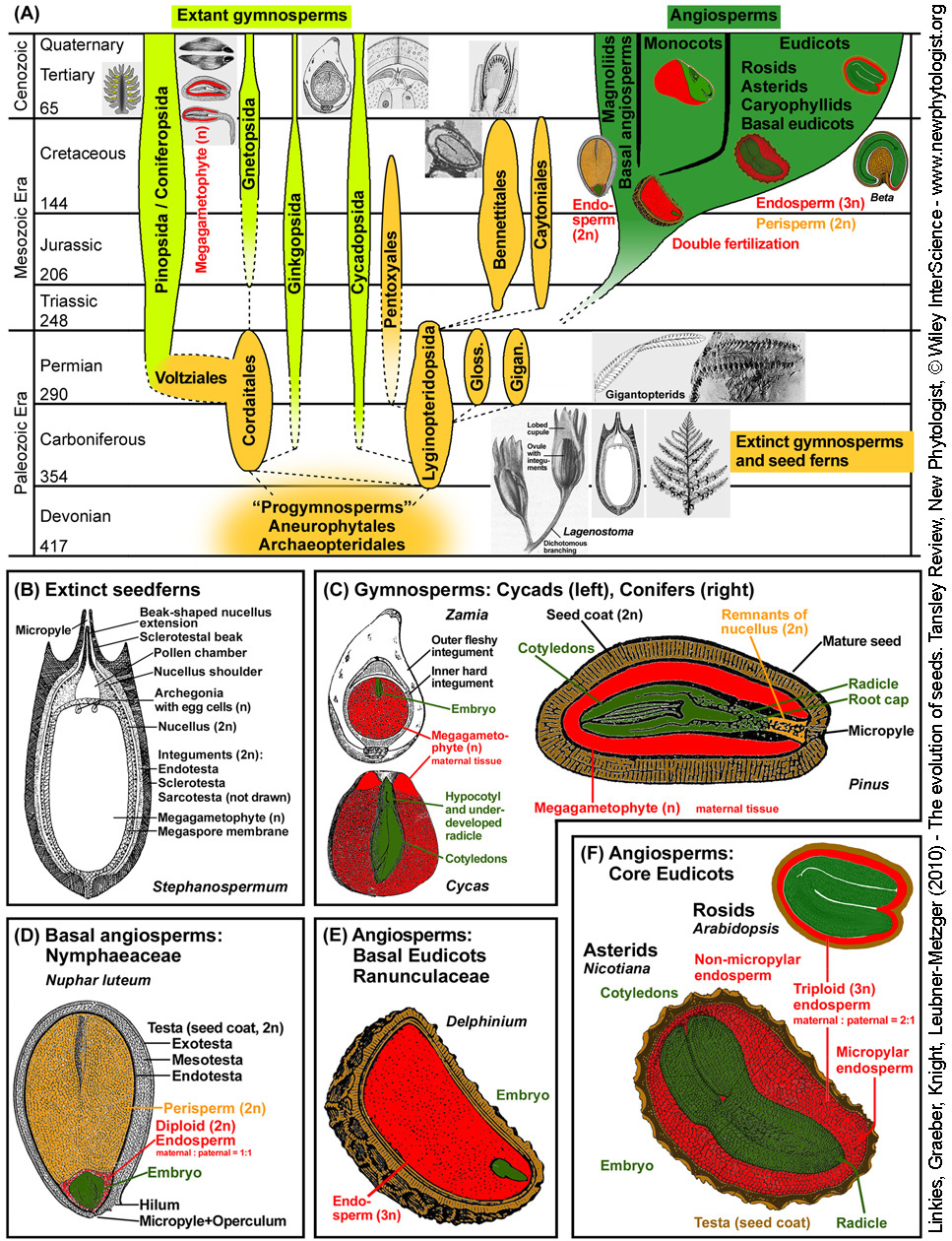
Figure 1. Origin and evolution of the seed habit.
(A) Seed plant phylogeny considering major extinct and extant gymnosperm and angiosperm clades. Based on molecular phylogenetic evidence the extant gymnosperms form a monophyletic group and the extant angiosperms form a distinct monophyletic group. Note that the precise evolutionary connections between the different gymnosperm groups are unknown and that the ancestors of the angiosperms are unknown. Extinct gymnosperm groups (insets of fossil drawings and images) include the paraphyletic group of seed ferns (Lyginopteridopsida), e.g. Devonian-Carboniferous Lyginopterids (e.g. Lagenostoma, from Scott 1909) and Carboniferous-Permian Medullosans (e.g. Stephanospermum, see panel B). Other groups/insets: Bennettitales (cycadeoids, from Scott 1909 & Zimmermann 1930), Glossopteridales (glossopterids), Gigantopteridales (gigantoperids, seed-bearing leaflet from Li & Yao, 1983), Gnetopsida (gnetophytes: Ephedridae, Gnetidae, Welwitschiidae), monocots (maize grain), Caryophyllids (Beta; Hermann et al. 2007). Time scale: Geological eras, periods, time in million years ago.
(B-F) Structural biodiversity of gymnosperm and angiosperm seeds with special consideration of the covering layers.
(B) Stephanospermum akenioides, drawing of a fossil medullosan seed fern ovule (Permian-Carboniferous Lyginopteridopsida; ca. 1 cm long). Within the megagametophyte archegonia with egg cells are evident. The megasporangium (nucellus) is surrounded by the integument which evolved into the three-layered testa. The micropylar extension, which in other Stephanospermum speciman form an apical funnel to capture wind-blown pollen, and a pollen chamber is evident. The nucellus and the megagametophyte are poorly perserved in the Stephanospermum ovule fossils. The megaspore membrane is robust and consists of a distinctive network of granules and rods of sporopollenin covered by a homogeneous outer layer; from Schnarf (1937).
(C) Mature seeds of extant gymnosperms (cycad and conifers). The diploid embryo is enveloped by the haploid megagametophyte and the diploid testa, remnants of the nucellus may be present. Cycad seeds (left): In the cycad ovule the micropylar opening produces a liquid pollination drop which catches wind-blown pollen and allows it to enter the pollen chamber. The pollen germinate and the pollen tubes grow into the nucellus tissue. There they release spermatozoids that swim to the archegonia and fertilize the egg cells. About half a year time difference is often found between cycad pollination and fertilization. The Cycas ('sago palm') embryo grows within the seed, and germination can occur only as the embryo has reached a similar size as the seed (MD). The embryo usually has 2-4 fleshy cotyledons, and the radicle is usually not fully developed as a distinct organ at the time of seed maturity. Seeds of tropical and subtropical Zamia species require many months of warm stratification before they will germinate (MPD). Conifer seed (right): Conifer ovules do not have pronounced pollen chambers and pollen grains do not release swimming spermazoids, but immobile sperms. The mature pine (Pinus spp.) seed contains an uncurved embryo with many cotyledons. The embryo is embedded in the megagametophyte tissue. The ovulate pine cone becomes woody as it matures. Seeds of Pinus spp. are either ND or have PD; from Engler (1926) and Schnarf (1937).
(D) Mature seed of a basal angiosperm (Nymphaeaceae, water lily) with diploid endosperm and perisperm; from Kirchner et al. 1938.
(E) Mature seed of a basal eudicot (Ranunculaceae) with abundant triploid endosperm and tiny embryo; from Engler & Prantl (1891).
(F) Mature seeds of core eudicots that differ in endosperm abundance: astrids (tobacco), rosids (Arabidopsis).
| Article in PDF format (900 KB) |
|
|
|
The Seed Biology Place |
Webdesign Gerhard Leubner 2000 |